
Biologia per il liceo/La genetica mendeliana
La genetica è lo studio dell'ereditarietà. Johann Gregor Mendel ha definito il quadro della genetica molto prima che i cromosomi o i geni fossero identificati, in un'epoca in cui la meiosi non era ben compresa. Mendel ha selezionato un semplice sistema biologico e ha condotto analisi quantitative metodiche utilizzando grandi dimensioni del campione. Grazie al lavoro di Mendel, sono stati rivelati i principi fondamentali dell'ereditarietà. Ora sappiamo che i geni, trasportati sui cromosomi, sono le unità funzionali di base dell'ereditarietà con la capacità di essere replicati, espressi o mutati. Oggi, i postulati avanzati da Mendel costituiscono la base della genetica classica o mendeliana. Non tutti i tratti vengono trasmessi dai genitori alla prole secondo la genetica mendeliana, ma gli esperimenti di Mendel servono come un eccellente punto di partenza per pensare all'ereditarietà.

Johann Gregor Mendel (1822–1884) fu uno studente, insegnante, scienziato e uomo di fede per tutta la vita. Da giovane adulto, entrò nell'abbazia agostiniana di San Tommaso a Brno, in quella che oggi è la Repubblica Ceca. Grazie al sostegno del monastero, insegnò fisica, botanica e corsi di scienze naturali a livello secondario e universitario. Nel 1856, iniziò un decennio di ricerca sui modelli di ereditarietà nelle api e nelle piante, scegliendo infine le piante di pisello come suo sistema modello primario (un sistema con caratteristiche utili utilizzate per studiare uno specifico fenomeno biologico da applicare ad altri sistemi). Nel 1865, Mendel presentò i risultati dei suoi esperimenti con quasi 30.000 piante di pisello alla locale Natural History Society. Dimostrò che i tratti vengono trasmessi dai genitori alla prole indipendentemente da altri tratti e in modelli dominanti e recessivi. Nel 1866 pubblicò il suo lavoro, Esperimenti di ibridazione vegetale, 1 negli atti della Società di storia naturale di Brünn.
Il lavoro di Mendel passò praticamente inosservato alla comunità scientifica, che credeva, erroneamente, che il processo di ereditarietà comportasse una fusione di tratti parentali che producevano un aspetto fisico intermedio nella prole. La teoria della fusione dell'ereditarietà affermava che i tratti parentali originali venivano persi o assorbiti dalla fusione nella prole, ma ora sappiamo che non è così. Questo processo ipotetico sembrava essere corretto a causa di ciò che ora conosciamo come variazione continua. La variazione continua deriva dall'azione di molti geni per determinare una caratteristica come l'altezza umana. La prole sembra essere una "fusione" dei tratti dei genitori.
Invece di caratteristiche continue, Mendel lavorò con tratti che venivano ereditati in classi distinte (in particolare, fiori viola rispetto a fiori bianchi); questo è definito variazione discontinua . La scelta di Mendel di questi tipi di tratti gli permise di vedere sperimentalmente che i tratti non venivano mescolati nella prole, né venivano assorbiti, ma piuttosto che mantenevano la loro distinzione e potevano essere trasmessi. Nel 1868, Mendel divenne abate del monastero e scambiò le sue attività scientifiche con i suoi doveri pastorali. Non fu riconosciuto per i suoi straordinari contributi scientifici durante la sua vita. Infatti, fu solo nel 1900 che il suo lavoro fu riscoperto, riprodotto e rivitalizzato da scienziati sul punto di scoprire la base cromosomica dell'ereditarietà.
Il modello di Mendel
[modifica | modifica sorgente]Il lavoro fondamentale di Mendel fu compiuto utilizzando il pisello da giardino, Pisum sativum, per studiare l'ereditarietà. Questa specie si autofeconda naturalmente, in modo che il polline incontri gli ovuli all'interno dei singoli fiori. I petali dei fiori rimangono sigillati ermeticamente fino a dopo l'impollinazione, impedendo l'impollinazione da parte di altre piante. Il risultato sono piante di pisello altamente consanguinee, o "di razza pura". Si tratta di piante che producono sempre una prole che assomiglia alla madre. Sperimentando con piante di pisello di razza pura, Mendel evitò la comparsa di tratti inaspettati nella prole che potrebbero verificarsi se le piante non fossero di razza pura. Il pisello da giardino cresce anche fino alla maturità in una stagione, il che significa che diverse generazioni potrebbero essere valutate in un tempo relativamente breve. Infine, grandi quantità di piselli da giardino potrebbero essere coltivate simultaneamente, consentendo a Mendel di concludere che i suoi risultati non erano semplicemente dovuti al caso.
-
 Pisum sativum
Pisum sativum -
 Pisum sativum
Pisum sativum -
 Pisum sativum
Pisum sativum -
 Il frutto di Pisum sativum
Il frutto di Pisum sativum -
 Caratteristiche che Mendel utilizzò per la sua ricerca
Caratteristiche che Mendel utilizzò per la sua ricerca





Gli incroci di Mendel
[modifica | modifica sorgente]Mendel eseguì delle ibridazioni , che comportano l'accoppiamento di due individui di razza pura con caratteristiche diverse. Nel pisello, che è naturalmente autoimpollinante, ciò avviene trasferendo manualmente il polline dall'antera di una pianta di pisello matura di una varietà allo stigma di una pianta di pisello matura separata della seconda varietà. Nelle piante, il polline trasporta i gameti maschili (sperma) allo stigma, un organo appiccicoso che intrappola il polline e consente allo sperma di muoversi lungo il pistillo fino ai gameti femminili (ovuli) sottostanti. Per impedire che la pianta di pisello che stava ricevendo il polline si autofecondasse e confondesse i suoi risultati, Mendel rimosse meticolosamente tutte le antere dai fiori della pianta prima che avessero la possibilità di maturare.
Per meglio capire il meccanismo di trasmissione dei caratteri Mendel esegui inizialmente degli incroci monoibridi, ovvero che riguardavano un singolo carattere, ad es. il colore del fiore.
Le piante utilizzate negli incroci di prima generazione erano chiamate P0 , o generazione parentale ( Figura sotto). Dopo ogni incrocio, Mendel raccoglieva i semi appartenenti alle piante P0 e li faceva crescere la stagione successiva. Questa prole era chiamata F1 , o prima generazione filiale ( filiale = prole, figlia o figlio). Una volta che Mendel esaminò le caratteristiche nella generazione F1 delle piante, lasciò che si autofecondassero naturalmente. Quindi raccolse e fece crescere i semi dalle piante F1 per produrre la F2 , o seconda generazione filiale. Gli esperimenti di Mendel si estesero oltre la generazione F2 alle generazioni F3 e F4 e così via, ma fu il rapporto delle caratteristiche nelle generazioni P0 − F1 −F2 a essere il più intrigante e a diventare la base per i postulati di Mendel.






Le caratteristiche dei piselli da giardino hanno rivelato le basi dell'ereditarietà
[modifica | modifica sorgente]Nella sua pubblicazione del 1865, Mendel riportò i risultati dei suoi incroci che comprendevano sette caratteristiche diverse, ciascuna con due tratti contrastanti. Un tratto è definito come una variazione nell'aspetto fisico di una caratteristica ereditaria. Le caratteristiche includevano altezza della pianta, consistenza del seme, colore del seme, colore del fiore, dimensione del baccello di pisello, colore del baccello di pisello e posizione del fiore. Per la caratteristica del colore del fiore, ad esempio, i due tratti contrastanti erano bianco contro viola. Per esaminare completamente ogni caratteristica, Mendel generò un gran numero di piante F1 e F2 , riportando i risultati di 19.959 piante F2 da sole. I suoi risultati erano coerenti.
Quali risultati trovò Mendel nei suoi incroci per il colore dei fiori? Innanzitutto, Mendel confermò di avere piante che si riproducevano correttamente per il colore dei fiori bianco o viola. Indipendentemente dal numero di generazioni esaminate da Mendel, tutta la prole autoincrociata di genitori con fiori bianchi aveva fiori bianchi e tutta la prole autoincrociata di genitori con fiori viola aveva fiori viola. Inoltre, Mendel confermò che, a parte il colore dei fiori, le piante di pisello erano fisicamente identiche.
Una volta completate queste convalide, Mendel applicò il polline di una pianta con fiori viola allo stigma di una pianta con fiori bianchi. Dopo aver raccolto e seminato i semi risultanti da questo incrocio, Mendel scoprì che il 100 percento della generazione ibrida F1 aveva fiori viola . La saggezza convenzionale a quel tempo (la teoria della fusione) avrebbe previsto che i fiori ibridi fossero viola pallido o che le piante ibride avessero un numero uguale di fiori bianchi e viola. In altre parole, ci si aspettava che i tratti parentali contrastanti si fondessero nella prole. Invece, i risultati di Mendel dimostrarono che il tratto dei fiori bianchi nella generazione F1 era completamente scomparso.
È importante notare che Mendel non interruppe lì la sua sperimentazione. Permise alle piante F1 di autofecondarsi e scoprì che, delle piante di generazione F2 , 705 avevano fiori viola e 224 fiori bianchi. Questo era un rapporto di 3,15 fiori viola per un fiore bianco, o approssimativamente 3:1. Quando Mendel trasferiva il polline da una pianta con fiori viola allo stigma di una pianta con fiori bianchi e viceversa, otteneva circa lo stesso rapporto indipendentemente da quale genitore, maschio o femmina, contribuisse a quale caratteristica. Questo è chiamato incrocio reciproco , un incrocio a coppie in cui le rispettive caratteristiche del maschio e della femmina in un incrocio diventano le rispettive caratteristiche della femmina e del maschio nell'altro incrocio. Per le altre sei caratteristiche esaminate da Mendel, le generazioni F1 e F2 si comportavano nello stesso modo in cui avevano fatto per il colore dei fiori. Una delle due caratteristiche sarebbe scomparsa completamente dalla generazione F1 solo per riapparire nella generazione F2 con un rapporto di approssimativamente 3:1.
I risultati delle ibridazioni dei piselli da giardino di Mendel
| Caratteristica | Contrasto dei tratti P0 | F1 Caratteristiche della prole | F2 Caratteristiche della prole | Rapporti di caratteristica F2 |
|---|---|---|---|---|
| Colore del fiore | Viola contro bianco | 100 percento viola |
|
3.15:1 |
| Posizione del fiore | Assiale vs. terminale | 100 percento assiale |
|
3.14:1 |
| Altezza della pianta | Alto contro nano | 100 percento alto |
|
2.84:1 |
| Tessitura del seme | Rotondo vs. rugoso | 100 percento rotondo |
|
2.96:1 |
| Colore del seme | Giallo contro verde | 100 percento giallo |
|
3.01:1 |
| Consistenza del baccello di pisello | Gonfiato vs. ristretto | 100 percento gonfiato |
|
2.95:1 |
| Colore del baccello di pisello | Verde contro giallo | 100% verde |
|
2.82:1 |
Dopo aver compilato i suoi risultati per molte migliaia di piante, Mendel concluse che le caratteristiche potevano essere divise in tratti espressi e latenti. Li chiamò rispettivamente tratti dominanti e recessivi. I tratti dominanti sono quelli che vengono ereditati senza modifiche in un'ibridazione. I tratti recessivi diventano latenti, o scompaiono, nella prole di un'ibridazione. Il tratto recessivo, tuttavia, riappare nella progenie della prole ibrida. Un esempio di tratto dominante è il tratto del fiore viola. Per questa stessa caratteristica (colore del fiore), i fiori di colore bianco sono un tratto recessivo. Il fatto che il tratto recessivo riapparisse nella generazione F2 significava che i tratti rimanevano separati (non mescolati) nelle piante della generazione F1. Mendel propose anche che le piante possedessero due copie del tratto per la caratteristica del colore del fiore e che ogni genitore trasmettesse una delle sue due copie alla sua prole, dove si uniscono. Inoltre, l'osservazione fisica di un tratto dominante potrebbe significare che la composizione genetica dell'organismo includeva due versioni dominanti della caratteristica o che includeva una versione dominante e una recessiva. Al contrario, l'osservazione di un tratto recessivo significava che l'organismo era privo di versioni dominanti di questa caratteristica.
Allora perché Mendel ottenne ripetutamente rapporti 3:1 nei suoi incroci? Per capire come Mendel dedusse i meccanismi basilari dell'ereditarietà che portano a tali rapporti, dobbiamo prima rivedere le leggi della probabilità.
Nozioni di base sulla probabilità
[modifica | modifica sorgente]Le probabilità sono misure matematiche di verosimiglianza. La probabilità empirica di un evento si calcola dividendo il numero di volte in cui l'evento si verifica per il numero totale di opportunità che l'evento si verifichi. È anche possibile calcolare probabilità teoriche dividendo il numero di volte in cui ci si aspetta che un evento si verifichi per il numero di volte in cui potrebbe verificarsi. Le probabilità empiriche derivano da osservazioni, come quelle di Mendel. Le probabilità teoriche, d'altro canto, derivano dalla conoscenza di come vengono prodotti gli eventi e dall'assunzione che le probabilità dei singoli risultati siano uguali. Una probabilità di uno per un evento indica che è garantito che si verifichi, mentre una probabilità di zero indica che è garantito che non si verifichi. Un esempio di evento genetico è un seme rotondo prodotto da una pianta di pisello.
In un esperimento, Mendel dimostrò che la probabilità che si verificasse l'evento "seme rotondo" era pari a uno nella prole F1 di genitori di razza pura, uno dei quali ha semi rotondi e uno dei quali ha semi rugosi. Quando le piante F1 furono successivamente auto-incrociate, la probabilità che una qualsiasi prole F2 avesse semi rotondi era ora di tre su quattro. In altre parole, in una grande popolazione di prole F2 scelta a caso, ci si aspettava che il 75 percento avesse semi rotondi, mentre ci si aspettava che il 25 percento avesse semi rugosi. Utilizzando un gran numero di incroci, Mendel fu in grado di calcolare le probabilità e di usarle per predire i risultati di altri incroci.
La regola del prodotto e la regola della somma
[modifica | modifica sorgente]Mendel dimostrò che le piante di pisello trasmettono le caratteristiche come unità discrete dal genitore alla prole. Come verrà discusso, Mendel determinò anche che diverse caratteristiche, come il colore e la consistenza dei semi, venivano trasmesse indipendentemente l'una dall'altra e potevano essere considerate in analisi di probabilità separate. Ad esempio, eseguendo un incrocio tra una pianta con semi verdi e rugosi e una pianta con semi gialli e rotondi si produceva comunque una prole che aveva un rapporto di 3:1 tra semi gialli/verdi (ignorando la consistenza dei semi) e un rapporto di 3:1 tra semi rugosi/rotondi (ignorando il colore dei semi). Le caratteristiche di colore e consistenza non si influenzavano a vicenda.
Regola del prodotto
[modifica | modifica sorgente]La regola del prodotto della probabilità può essere applicata a questo fenomeno della trasmissione indipendente delle caratteristiche. La regola del prodotto afferma che la probabilità che due eventi indipendenti si verifichino insieme può essere calcolata moltiplicando le probabilità individuali di ciascun evento che si verifica da solo. Per dimostrare la regola del prodotto, immagina di lanciare un dado a sei facce (D) e di lanciare una moneta da un centesimo (P) contemporaneamente. Il dado può lanciare qualsiasi numero da 1 a 6 (D # ), mentre la moneta da un centesimo può girare testa (PT) o croce (PC) . L'esito del lancio del dado non ha effetto sull'esito del lancio della moneta da un centesimo e viceversa. Ci sono 12 possibili esiti di questa azione ( Tabella sotto) e ci si aspetta che ogni evento si verifichi con la stessa probabilità.
Dodici risultati ugualmente probabili del lancio di un dado e del lancio di un penny
| Dado rotante | Penny che gira |
|---|---|
| D1 | PT |
| D1 | PC |
| D2 | PT |
| D2 | PC |
| D3 | PT |
| D3 | PC |
| D4 | PT |
| D4 | PC |
| D5 | PT |
| D5 | PC |
| D6 | PT |
| D6 | PC |
Dei 12 possibili risultati, il dado ha una probabilità di 2/12 (o 1/6) di ottenere un due, e il penny ha una probabilità di 6/12 (o 1/2) di ottenere testa. In base alla regola del prodotto, la probabilità di ottenere il risultato combinato 2 e testa è: (D 2 ) x (P H ) = (1/6) x (1/2) o 1/12. Nota la parola "e" nella descrizione della probabilità. "e" è un segnale per applicare la regola del prodotto. Ad esempio, considera come la regola del prodotto viene applicata all'incrocio diibrido: la probabilità di avere entrambi i tratti dominanti (ad esempio, giallo e rotondo) nella progenie F2 è il prodotto delle probabilità di avere il tratto dominante per ciascuna caratteristica, come mostrato qui:
3/4 × 3/4 = 9/16
Regola della somma
[modifica | modifica sorgente]D'altro canto, la regola della somma delle probabilità viene applicata quando si considerano due risultati reciprocamente esclusivi che possono verificarsi tramite più di un percorso. La regola della somma afferma che la probabilità del verificarsi di un evento o dell'altro evento, di due eventi reciprocamente esclusivi, è la somma delle loro probabilità individuali. Nota la parola "o" nella descrizione della probabilità. "o" indica che dovresti applicare la regola della somma. In questo caso, immaginiamo di lanciare un penny (P) e un quarto (Q). Qual è la probabilità che una moneta esca testa e l'altra croce? Questo risultato può essere ottenuto in due casi: il penny può essere testa (PH ) e il quarto può essere croce (QT ) , oppure il quarto può essere testa (QH ) e il penny può essere croce (PT ) . Entrambi i casi soddisfano il risultato. Con la regola della somma, calcoliamo la probabilità di ottenere una testa e una croce come [(PH ) × (QT ) ] + [(QH ) × (PT ) ] = [(1/2) × (1/2)] + [(1/2) × (1/2)] = 1/2 ( Tabella sotto). Dovresti anche notare che abbiamo usato la regola del prodotto per calcolare la probabilità di PH e QT , e anche la probabilità di PT e QH , prima di sommarle. Di nuovo, la regola della somma può essere applicata per mostrare la probabilità di avere esattamente un tratto dominante nella generazione F2 di un incrocio diibrido:
La regola del prodotto e la regola della somma
| Regola del prodotto | Regola della somma |
|---|---|
| Per gli eventi indipendenti A e B, la probabilità (P) che si verifichino entrambi (A e B) è (P A × P B ) | Per gli eventi mutuamente esclusivi A e B, la probabilità (P) che si verifichi almeno uno (A o B) è (P A + P B ) |
Per usare le leggi di probabilità nella pratica, dobbiamo lavorare con grandi dimensioni del campione perché piccole dimensioni del campione sono soggette a deviazioni causate dal caso. Le grandi quantità di piante di pisello che Mendel esaminò gli permisero di calcolare le probabilità dei tratti che apparivano nella sua generazione F2. Come imparerai, questa scoperta significava che quando i tratti parentali erano noti, i tratti della prole potevano essere previsti con precisione anche prima della fecondazione.
La genetica mendeliana moderna
[modifica | modifica sorgente]Le caratteristiche fisiche sono espresse tramite geni (che Mendel chiamava "fattori") trasportati sui cromosomi. Per ogni gene esistono due varianti simili o uguali, omologhi, presenti sui rispettivi cromosomi omologhi. Ogni coppia di cromosomi omologhi ha lo stesso ordine lineare di geni omologhi (alleli). I piselli sono organismi diploidi in quanto hanno due copie di ogni cromosoma. Lo stesso vale per molte altre piante e per praticamente tutti gli animali. Gli organismi diploidi producono gameti aploidi, che contengono una copia di ogni cromosoma omologo che si unisce alla fecondazione per creare uno zigote diploide.
Nei casi in cui un singolo gene controlla una singola caratteristica, un organismo diploide ha due copie di geni (omologhi) che possono o meno codificare la stessa versione di quella caratteristica. Le varianti geniche che derivano da mutazione ed esistono nelle stesse posizioni relative sui cromosomi omologhi sono chiamate alleli . Mendel ha esaminato l'ereditarietà dei geni con solo due forme alleliche, ma è comune incontrare più di due alleli per qualsiasi gene dato in una popolazione naturale.
-
 Cromosoma con evidenziato un gene
Cromosoma con evidenziato un gene -
 Cromosoma duplicato
Cromosoma duplicato -
 Cromosoma Y umano (non duplicato) in cui si evidenzia il gene Sry
Cromosoma Y umano (non duplicato) in cui si evidenzia il gene Sry -
 Cromosomi omologhi
Cromosomi omologhi -
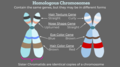 Cromosomi omologhi e alleli
Cromosomi omologhi e alleli





Fenotipi e genotipi
[modifica | modifica sorgente]Due alleli per un dato gene in un organismo diploide vengono espressi e interagiscono per produrre caratteristiche fisiche. I tratti osservabili espressi da un organismo sono indicati come fenotipo . Il corredo genetico sottostante di un organismo è chiamato genotipo, che corrisponde quindi alla coppia di alleli . Gli esperimenti di ibridazione di Mendel dimostrano la differenza tra fenotipo e genotipo. Quando piante di razza pura in cui un genitore aveva baccelli gialli e uno aveva baccelli verdi venivano fecondate in modo incrociato, tutta la prole ibrida F1 aveva baccelli gialli. Vale a dire, la prole ibrida era fenotipicamente identica al genitore di razza pura con baccelli gialli. Tuttavia, sappiamo che l'allele donato dal genitore con baccelli verdi non è stato semplicemente perso perché è ricomparso in alcuni dei discendenti F2. Pertanto, le piante F1 devono essere state genotipicamente diverse dal genitore con baccelli gialli.
Se i due alleli relativi ad un certo carattere (genotipo) sono uguali l'individuo viene detto omozigote per quel carattere; se sono diversi viene detto eterozigote. Le piante P1 che Mendel usò nei suoi esperimenti erano tutte omozigoti per il tratto che stava studiando. Gli organismi diploidi che sono omozigoti per un dato gene, o locus, hanno due alleli identici per quel gene sui loro cromosomi omologhi. Le piante parentali di pisello di Mendel si riproducevano sempre correttamente perché entrambi i gameti prodotti portavano lo stesso tratto. Quando le piante P1 con tratti diversi venivano fecondate in modo incrociato, tutta la prole era eterozigote per il tratto diverso, il che significa che il loro genotipo era formato da alleli diversi per il gene esaminato.
-
 Genotipo e fenotipo
Genotipo e fenotipo -
 Genotipo e fenotipo
Genotipo e fenotipo -
 Il genotipo, influenzato dall'ambiente, produce il fenotipo
Il genotipo, influenzato dall'ambiente, produce il fenotipo -
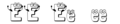 Il genotipo si rappresenta con una coppia di lettere: maiuscolo se dominante, minuscolo se recessivo
Il genotipo si rappresenta con una coppia di lettere: maiuscolo se dominante, minuscolo se recessivo



Alleli dominanti e recessivi
[modifica | modifica sorgente]La nostra discussione sugli organismi omozigoti ed eterozigoti ci porta al motivo per cui la prole eterozigote F1 era identica a uno dei genitori, anziché esprimere entrambi gli alleli. In tutte e sette le caratteristiche della pianta di pisello, uno dei due alleli contrastanti era dominante e l'altro era recessivo. Mendel chiamò l'allele dominante fattore unitario espresso; l'allele recessivo era definito fattore unitario latente. Ora sappiamo che questi cosiddetti fattori unitari sono in realtà geni su coppie di cromosomi omologhi. Per un gene espresso in uno schema dominante e recessivo, gli organismi omozigoti dominanti ed eterozigoti appariranno identici (ovvero avranno genotipi diversi ma lo stesso fenotipo). I tratti dell'allele recessivo saranno osservati solo negli individui omozigoti recessivi ( Tabella sotto).
Ereditarietà umana in modelli dominanti e recessivi
| Tratti dominanti | Tratti recessivi | |||
|---|---|---|---|---|
| Acondroplasia |  |
Albinismo |  | |
| Brachidattilia |  |
Fibrosi cistica |  | |
| Malattia di Huntington |  |
Distrofia muscolare di Duchenne |  | |
| Sindrome di Marfan |  |
Galattosemia |  | |
| Neurofibromatosi |  |
Fenilchetonuria |  | |
| Picco della vedova |  |
Anemia falciforme |  | |
| Capelli lanosi |  |
Malattia di Tay-Sachs |  | |
Esistono diverse convenzioni per fare riferimento a geni e alleli. Ai fini di questo capitolo, abbrevieremo i geni utilizzando la prima lettera del tratto dominante corrispondente al gene. Ad esempio, il viola è il tratto dominante per il colore dei fiori di una pianta di pisello, quindi il gene del colore dei fiori sarebbe abbreviato come V (si noti che è consuetudine mettere in corsivo le designazioni dei geni). Inoltre, useremo lettere maiuscole e minuscole per rappresentare rispettivamente gli alleli dominanti e recessivi. Pertanto, faremmo riferimento al genotipo di una pianta di pisello omozigote dominante con fiori viola come VV , una pianta di pisello omozigote recessiva con fiori bianchi come vv e una pianta di pisello eterozigote con fiori viola come Vv .
L'approccio del quadrato di Punnett per un incrocio monoibrido
[modifica | modifica sorgente]Quando la fecondazione avviene tra due genitori di razza pura che differiscono solo per una caratteristica, il processo è chiamato incrocio monoibrido e la prole risultante è monoibrida. Mendel ha eseguito sette incroci monoibridi che coinvolgono tratti contrastanti per ogni caratteristica. Sulla base dei suoi risultati nelle generazioni F1 e F2 , Mendel ha postulato che ogni genitore nell'incrocio monoibrido ha contribuito con uno dei due fattori unitari accoppiati a ogni prole e che ogni possibile combinazione di fattori unitari era ugualmente probabile.
Per dimostrare un incrocio monoibrido, si consideri il caso di piante di pisello di razza pura con semi di pisello gialli rispetto a quelli verdi. Il colore dominante dei semi è il giallo; pertanto, i genotipi parentali erano rispettivamente YY per le piante con semi gialli e yy per le piante con semi verdi. È possibile disegnare un quadrato di Punnett , ideato dal genetista britannico Reginald Punnett, che applica le regole della probabilità per predire i possibili risultati di un incrocio o accoppiamento genetico e le loro frequenze attese. Per preparare un quadrato di Punnett, tutte le possibili combinazioni degli alleli parentali vengono elencate lungo la parte superiore (per un genitore) e laterale (per l'altro genitore) di una griglia, che rappresenta la loro segregazione meiotica in gameti aploidi. Quindi le combinazioni di uovo e sperma vengono create nelle caselle della tabella per mostrare quali alleli si stanno combinando. Ogni casella rappresenta quindi il genotipo diploide di uno zigote, o uovo fecondato, che potrebbe derivare da questo accoppiamento. Poiché ogni possibilità è ugualmente probabile, i rapporti genotipici possono essere determinati da un quadrato di Punnett. Se è noto il modello di ereditarietà (dominante o recessivo), è possibile dedurre anche i rapporti fenotipici. Per un incrocio monoibrido di due genitori di razza pura, ogni genitore contribuisce con un tipo di allele. In questo caso, è possibile un solo genotipo. Tutta la prole è Yy e ha semi gialli ( Figura sotto ).

Un autoincrocio di una delle prole eterozigote Yy può essere rappresentato in un quadrato di Punnett 2 × 2 perché ogni genitore può donare uno di due diversi alleli. Pertanto, la prole può potenzialmente avere una delle quattro combinazioni di alleli: YY , Yy , yY o yy ( Figura sopra). Si noti che ci sono due modi per ottenere il genotipo Yy : una Y dall'uovo e una y dallo sperma, oppure una y dall'uovo e una Y dallo sperma. Entrambe queste possibilità devono essere contate. Si ricordi che le caratteristiche della pianta di pisello di Mendel si comportavano allo stesso modo negli incroci reciproci. Pertanto, le due possibili combinazioni eterozigote producono una prole che è genotipicamente e fenotipicamente identica nonostante i loro alleli dominanti e recessivi derivino da genitori diversi. Sono raggruppati insieme. Poiché la fecondazione è un evento casuale, ci aspettiamo che ogni combinazione sia ugualmente probabile e che la prole mostri un rapporto di genotipi YY : Yy : yy di 1:2:1. Inoltre, poiché la prole YY e Yy ha semi gialli e sono fenotipicamente identici, applicando la regola della somma delle probabilità, ci aspettiamo che la prole mostri un rapporto fenotipico di 3 giallo:1 verde. Infatti, lavorando con grandi dimensioni del campione, Mendel ha osservato approssimativamente questo rapporto in ogni generazione F2 risultante da incroci per tratti individuali.
Mendel ha convalidato questi risultati eseguendo un incrocio F 3 in cui ha auto-incrociato le piante F2 che esprimevano il carattere dominante e recessivo. Quando ha auto-incrociato le piante che esprimevano semi verdi, tutta la prole aveva semi verdi, confermando che tutti i semi verdi avevano genotipi omozigoti di yy . Quando ha auto-incrociato le piante F2 che esprimevano semi gialli, ha scoperto che un terzo delle piante si riproduceva in modo vero e due terzi delle piante si segregavano con un rapporto di 3:1 di semi gialli:verdi. In questo caso, le piante di riproduzione vera avevano genotipi omozigoti ( YY ), mentre le piante segreganti corrispondevano al genotipo eterozigote ( Yy ). Quando queste piante si autofecondavano, il risultato era esattamente come l' incrocio autofecondante F1 .
Il test cross distingue il fenotipo dominante
[modifica | modifica sorgente]Oltre a predire la prole di un incrocio tra genitori omozigoti o eterozigoti noti, Mendel sviluppò anche un modo per determinare se un organismo che esprimeva un tratto dominante fosse eterozigote o omozigote. Chiamata test-cross , questa tecnica è ancora utilizzata dagli allevatori di piante e animali. In un incrocio di prova, l'organismo che esprime il tratto dominante viene incrociato con un organismo che è omozigote recessivo per la stessa caratteristica. Se l'organismo che esprime il tratto dominante è omozigote, allora tutta la prole F1 sarà eterozigote che esprime il tratto dominante ( Figura 12.5 ). In alternativa, se l'organismo che esprime il tratto dominante è eterozigote, la prole F1 mostrerà un rapporto 1:1 di eterozigoti e omozigoti recessivi ( Figura 12.5 ). L'incrocio di prova convalida ulteriormente il postulato di Mendel secondo cui le coppie di fattori unitari si segregano equamente.

Esercizio: nelle piante di pisello, i piselli rotondi ( R ) sono dominanti sui piselli rugosi ( r ). Si esegue un incrocio di prova tra una pianta di pisello con piselli rugosi (genotipo rr ) e una pianta di genotipo sconosciuto che ha piselli rotondi. Si ottengono tre piante, tutte con piselli rotondi. Da questi dati, si può dire se la pianta madre del pisello rotondo è omozigote dominante o eterozigote? Se la pianta madre del pisello rotondo è eterozigote, qual è la probabilità che un campione casuale di 3 piselli di progenie siano tutti rotondi?
Molte malattie umane sono ereditate geneticamente. Una persona sana in una famiglia in cui alcuni membri soffrono di un disturbo genetico recessivo potrebbe voler sapere se ha il gene che causa la malattia e quale rischio esiste di trasmettere il disturbo alla prole. Naturalmente, fare un test cross negli esseri umani è immorale e poco pratico. Invece, i genetisti usano l' analisi del pedigree per studiare il modello di ereditarietà delle malattie genetiche umane ( Figura sotto).

Figura sopra. L'alcaptonuria è un disturbo genetico recessivo in cui due aminoacidi, fenilalanina e tirosina, non vengono metabolizzati correttamente. Gli individui affetti possono avere la pelle scura e l'urina marrone e possono soffrire di danni alle articolazioni e altre complicazioni. In questo pedigree, gli individui affetti dal disturbo sono indicati in blu e hanno il genotipo aa . Gli individui non affetti sono indicati in giallo e hanno il genotipo AA o Aa . Si noti che è spesso possibile determinare il genotipo di una persona dal genotipo della sua prole. Ad esempio, se nessuno dei genitori ha il disturbo ma il figlio sì, devono essere eterozigoti. Due individui nel pedigree hanno un fenotipo non affetto ma un genotipo sconosciuto. Poiché non hanno il disturbo, devono avere almeno un allele normale, quindi il loro genotipo ottiene la designazione " A? ". Quali sono i genotipi degli individui etichettati 1, 2 e 3?
Letalità
[modifica | modifica sorgente]Una grande percentuale di geni nel genoma di un individuo è essenziale per la sopravvivenza. Occasionalmente, un allele non funzionale per un gene essenziale può sorgere tramite mutazione ed essere trasmesso in una popolazione, purché gli individui con questo allele abbiano anche una copia funzionale di tipo selvatico. L'allele di tipo selvatico funziona a una capacità sufficiente a sostenere la vita ed è quindi considerato dominante sull'allele non funzionale. Tuttavia, considera due genitori eterozigoti che hanno un genotipo di tipo selvatico/mutante non funzionale per un ipotetico gene essenziale. In un quarto della loro prole, ci aspetteremmo di osservare individui che sono omozigoti recessivi per l'allele non funzionale. Poiché il gene è essenziale, questi individui potrebbero non svilupparsi dopo la fecondazione, morire in utero o morire più tardi nella vita, a seconda di quale fase della vita richiede questo gene. Un modello di ereditarietà in cui un allele è letale solo nella forma omozigote e in cui l'eterozigote può essere normale o avere un fenotipo alterato non letale è definito letale recessivo .
Per gli incroci tra individui eterozigoti con un allele letale recessivo che causa la morte prima della nascita quando omozigoti, si osserverebbero solo omozigoti ed eterozigoti di tipo selvatico. Il rapporto genotipico sarebbe quindi 2:1. In altri casi, l'allele letale recessivo potrebbe anche presentare un fenotipo dominante (ma non letale) nell'eterozigote. Ad esempio, l'allele letale recessivo Curly in Drosophila influenza la forma delle ali nella forma eterozigote ma è letale nell'omozigote.

Una singola copia dell'allele di tipo selvatico non è sempre sufficiente per il normale funzionamento o persino per la sopravvivenza. Il modello di ereditarietà letale dominante è quello in cui un allele è letale sia nell'omozigote che nell'eterozigote; questo allele può essere trasmesso solo se il fenotipo di letalità si verifica dopo l'età riproduttiva. Gli individui con mutazioni che determinano alleli letali dominanti non riescono a sopravvivere nemmeno nella forma eterozigote. Gli alleli letali dominanti sono molto rari perché, come ci si potrebbe aspettare, l'allele dura solo una generazione e non viene trasmesso. Tuttavia, proprio come l'allele letale recessivo potrebbe non manifestare immediatamente il fenotipo di morte, anche gli alleli letali dominanti potrebbero non essere espressi fino all'età adulta. Una volta che l'individuo raggiunge l'età riproduttiva, l'allele potrebbe essere trasmesso inconsapevolmente, con conseguente morte ritardata in entrambe le generazioni. Un esempio di ciò negli esseri umani è la malattia di Huntington, in cui il sistema nervoso si consuma gradualmente ( Figura sopra). Le persone eterozigoti per l'allele dominante di Huntington ( Hh ) svilupperanno inevitabilmente la malattia mortale. Tuttavia, l'insorgenza della malattia di Huntington potrebbe non verificarsi prima dei 40 anni, momento in cui le persone affette potrebbero aver già trasmesso l'allele al 50 percento della loro prole
La legge della dominanza: gli alleli possono essere dominanti o recessivi
[modifica | modifica sorgente]La legge di dominanza di Mendel afferma che in un eterozigote, un tratto nasconderà la presenza di un altro tratto per la stessa caratteristica. Invece di entrambi gli alleli che contribuiscono a un fenotipo, l'allele dominante sarà espresso esclusivamente. L'allele recessivo rimarrà "latente" ma sarà trasmesso alla prole nello stesso modo in cui viene trasmesso l'allele dominante. Il tratto recessivo sarà espresso solo dalla prole che ha due copie di questo allele ( Figura sotto), e questa prole si riprodurrà in modo fedele quando auto-incrociata.
Dagli esperimenti di Mendel con le piante di pisello, i ricercatori hanno scoperto che la legge della dominanza non è sempre vera. Invece, sono stati scoperti diversi modelli di ereditarietà.

La legge della segregazione
[modifica | modifica sorgente]Osservando che le piante di pisello di razza pura con tratti contrastanti davano origine a generazioni F1 che esprimevano tutte il tratto dominante e a generazioni F2 che esprimevano i tratti dominanti e recessivi in un rapporto 3:1, Mendel propose la legge della segregazione . Questa legge afferma che i fattori unitari accoppiati (geni) devono segregarsi equamente nei gameti in modo che la prole abbia la stessa probabilità di ereditare entrambi i fattori. Per la generazione F2 di un incrocio monoibrido, potrebbero risultare le seguenti tre possibili combinazioni di genotipi: omozigote dominante, eterozigote o omozigote recessivo. Poiché gli eterozigoti potrebbero derivare da due percorsi diversi (ricevendo un allele dominante e uno recessivo da entrambi i genitori) e poiché gli eterozigoti e gli individui omozigoti dominanti sono fenotipicamente identici, la legge supporta il rapporto fenotipico 3:1 osservato da Mendel. La segregazione equa degli alleli è il motivo per cui possiamo applicare il quadrato di Punnett per predire accuratamente la prole di genitori con genotipi noti. La base fisica della legge di segregazione di Mendel è la prima divisione della meiosi, in cui i cromosomi omologhi con le loro diverse versioni di ogni gene vengono segregati in nuclei figli. Il ruolo della segregazione meiotica dei cromosomi nella riproduzione sessuale non era compreso dalla comunità scientifica durante la vita di Mendel.
La legge dell'assortimento indipendente
[modifica | modifica sorgente]La legge di Mendel sull'assortimento indipendente afferma che i geni non si influenzano a vicenda per quanto riguarda la classificazione degli alleli in gameti e che ogni possibile combinazione di alleli per ogni gene ha la stessa probabilità di verificarsi. L'assortimento indipendente dei geni può essere illustrato dall'incrocio diibrido , un incrocio tra due genitori di razza pura che esprimono tratti diversi per due caratteristiche. Considera le caratteristiche del colore e della consistenza dei semi per due piante di pisello, una con semi verdi e rugosi ( yyrr ) e un'altra con semi gialli e rotondi ( YYRR ). Poiché ogni genitore è omozigote, la legge di segregazione indica che i gameti per la pianta verde/rugosa sono tutti yr e i gameti per la pianta gialla/rotonda sono tutti YR . Pertanto, la generazione F1 della prole è tutta YyRr ( Figura sotto).

Nelle piante di pisello, la forma rotonda del seme (R) è dominante rispetto alla forma rugosa del seme (r) e i piselli gialli (Y) sono dominanti rispetto ai piselli verdi (y). Quali sono i possibili genotipi e fenotipi per un incrocio tra piante di pisello RrYY e rrYy? Di quanti quadrati hai bisogno per fare un'analisi del quadrato di Punnett di questo incrocio?
Per la generazione F2, la legge di segregazione richiede che ogni gamete riceva un allele R o un allele r insieme a un allele Y o un allele y . La legge dell'assortimento indipendente afferma che un gamete in cui è stato ordinato un allele r avrebbe la stessa probabilità di contenere un allele Y o un allele y . Quindi, ci sono quattro gameti ugualmente probabili che possono essere formati quando l' eterozigote YyRr viene auto-incrociato, come segue: YR , Yr , yR e yr . Disponendo questi gameti lungo la parte superiore e sinistra di un quadrato di Punnett 4 × 4 otteniamo 16 combinazioni genotipiche ugualmente probabili. Da questi genotipi, deduciamo un rapporto fenotipico di 9 rotondo/giallo:3 rotondo/verde:3 rugoso/giallo:1 rugoso/verde ( Figura 12.16 ). Questi sono i rapporti di prole che ci aspetteremmo, supponendo di aver effettuato gli incroci con un campione sufficientemente ampio.
A causa dell'assortimento e della dominanza indipendenti, il rapporto fenotipico diibrido 9:3:3:1 può essere ridotto a due rapporti 3:1, caratteristici di qualsiasi incrocio monoibrido che segue uno schema dominante e recessivo. Ignorando il colore del seme e considerando solo la consistenza del seme nell'incrocio diibrido di cui sopra, ci aspetteremmo che tre quarti della prole della generazione F2 siano rotondi e un quarto rugosi. Allo stesso modo, isolando solo il colore del seme, supporremmo che tre quarti della prole F2 siano gialli e un quarto verde. L'ordinamento degli alleli per consistenza e colore sono eventi indipendenti, quindi possiamo applicare la regola del prodotto. Pertanto, ci si aspetta che la proporzione di prole F2 rotonda e gialla sia (3/4) × (3/4) = 9/16 e che la proporzione di prole rugosa e verde sia (1/4) × (1/4) = 1/16. Queste proporzioni sono identiche a quelle ottenute utilizzando un quadrato di Punnett. Anche la prole rotonda, verde e rugosa, gialla può essere calcolata utilizzando la regola del prodotto, poiché ciascuno di questi genotipi include un fenotipo dominante e uno recessivo. Pertanto, la proporzione di ciascuno è calcolata come (3/4) × (1/4) = 3/16.
La legge dell'assortimento indipendente indica anche che un incrocio tra genitori gialli e rugosi ( YYrr ) e verdi e rotondi ( yyRR ) produrrebbe la stessa prole F1 e F2 dell'incrocio YYRR x yyrr .
La base fisica della legge dell'assortimento indipendente risiede anche nella meiosi I, in cui le diverse coppie omologhe si allineano in orientamenti casuali. Ogni gamete può contenere qualsiasi combinazione di cromosomi paterni e materni (e quindi i geni su di essi) perché l'orientamento delle tetradi sul piano della metafase è casuale.


Calcolare la probabilità di prole negli incroci multiibridi
[modifica | modifica sorgente]Metodo della biforcazione
[modifica | modifica sorgente]Quando si considerano più di due geni, il metodo del quadrato di Punnett diventa poco maneggevole. Ad esempio, esaminare un incrocio che coinvolge quattro geni richiederebbe una griglia 16 × 16 contenente 256 caselle. Sarebbe estremamente macchinoso immettere manualmente ogni genotipo. Per incroci più complessi, sono preferiti i metodi della biforcazione e della probabilità.
Per preparare un diagramma a linee biforcate per un incrocio tra eterozigoti F1 risultante da un incrocio tra genitori AABBCC e aabbcc , creiamo prima righe pari al numero di geni considerati, quindi segreghiamo gli alleli in ogni riga su linee biforcate in base alle probabilità per singoli incroci monoibridi ( Figura sotto). Quindi moltiplichiamo i valori lungo ogni percorso biforcato per ottenere le probabilità della prole F2 . Si noti che questo processo è una versione diagrammatica della regola del prodotto. I valori lungo ogni percorso biforcato possono essere moltiplicati perché ogni gene si assortisce in modo indipendente. Per un incrocio triibrido, il rapporto fenotipico F2 è 27:9:9:9:3:3:3:1.

Metodo di probabilità
[modifica | modifica sorgente]Mentre il metodo della linea biforcuta è un approccio diagrammatico per tenere traccia delle probabilità in un incrocio, il metodo della probabilità fornisce le proporzioni di prole che si prevede esibisca ogni fenotipo (o genotipo) senza l'assistenza visiva aggiuntiva. Entrambi i metodi utilizzano la regola del prodotto e considerano gli alleli per ogni gene separatamente. In precedenza, abbiamo esaminato le proporzioni fenotipiche per un incrocio triibrido utilizzando il metodo della linea biforcuta; ora utilizzeremo il metodo della probabilità per esaminare le proporzioni genotipiche per un incrocio con ancora più geni.
Per un incrocio triibrido, scrivere il metodo della linea biforcuta è noioso, anche se non quanto usare il metodo del quadrato di Punnett. Per dimostrare appieno la potenza del metodo di probabilità, tuttavia, possiamo considerare calcoli genetici specifici. Ad esempio, per un incrocio tetraibrido tra individui eterozigoti per tutti e quattro i geni, e in cui tutti e quattro i geni si ordinano indipendentemente e in uno schema dominante e recessivo, quale proporzione della prole ci si aspetterà che sia omozigote recessiva per tutti e quattro gli alleli? Invece di scrivere ogni possibile genotipo, possiamo usare il metodo di probabilità. Sappiamo che per ogni gene, la frazione di prole omozigote recessiva sarà 1/4. Pertanto, moltiplicando questa frazione per ciascuno dei quattro geni, (1/4) × (1/4) × (1/4) × (1/4), determiniamo che 1/256 della prole sarà quadruplicatamente omozigote recessiva.
Per lo stesso incrocio tetraibrido, qual è la proporzione prevista di prole che ha il fenotipo dominante in tutti e quattro i loci? Possiamo rispondere a questa domanda usando le proporzioni fenotipiche, ma facciamolo nel modo più difficile, usando le proporzioni genotipiche. La domanda chiede la proporzione di prole che è 1) omozigote dominante in A o eterozigote in A, e 2) omozigote in B o eterozigote in B , e così via. Annotare "o" e "e" in ogni circostanza chiarisce dove applicare le regole di somma e prodotto. La probabilità di un omozigote dominante in A è 1/4 e la probabilità di un eterozigote in A è 1/2. La probabilità dell'omozigote o dell'eterozigote è 1/4 + 1/2 = 3/4 usando la regola della somma. La stessa probabilità può essere ottenuta nello stesso modo per ciascuno degli altri geni, in modo che la probabilità di un fenotipo dominante in A e B e C e D sia, usando la regola del prodotto, uguale a 3/4 × 3/4 × 3/4 × 3/4, ovvero 81/256. Se non sei sicuro di come combinare le probabilità, tornare al metodo della linea biforcuta dovrebbe chiarirti le idee.
Regole per la fecondazione multiibrida
[modifica | modifica sorgente]Prevedere i genotipi e i fenotipi della prole da incroci dati è il modo migliore per testare la tua conoscenza della genetica mendeliana. Dato un incrocio multiibrido che obbedisce all'assortimento indipendente e segue uno schema dominante e recessivo, esistono diverse regole generalizzate; puoi usare queste regole per controllare i tuoi risultati mentre lavori sui calcoli genetici ( Tabella sotto). Per applicare queste regole, devi prima determinare n , il numero di coppie di geni eterozigoti (il numero di geni che segregano due alleli ciascuno). Ad esempio, un incrocio tra eterozigoti AaBb e AaBb ha un n di 2. Al contrario, un incrocio tra AABb e AABb ha un n di 1 perché A non è eterozigote.
Regole generali per gli incroci multiibridi
| Regola generale | Numero di coppie di geni eterozigoti |
|---|---|
| Numero di gameti F1 diversi | 2n |
| Numero di diversi genotipi F2 | 3n |
| Data l'eredità dominante e recessiva, il numero di diversi fenotipi F2 | 2n |
Attività
[modifica | modifica sorgente]Esercizi di genetica su Wikiversità
Esercitazione di genetica su Wikiversità
Esercizi sugli alberi genealogici in inglese (genetica mendeliana e post-mendeliana) (su LibreTexts da KanAcademy)
Diversi tipi di esercizi con diverse difficoltà, ordinati per categoria, di genetica in generale, con soluzioni
Esercizi sulla genetica mendeliana in inglese
Esercizi un po' più difficili e di riflessione in inglese